

GÉNÉTIQUE
En cours de corrections...
La génétique est la science qui étudie l'hérédité, c'est-à-dire la transmission de caractères héréditaires entre les géniteurs et leur descendance. Ici, nous parlerons principalement de la génétique des couleurs, intéressantes dans l'élevage.
Nul besoin de connaître la génétique pour avoir un Axolotl, et même en élever. Cependant, la génétique s'avère une alliée intéressante pour prévoir et organiser les pontes, pour comprendre comment une maladie génétique se transmet, etc.
Grâce à elle, on comprend pourquoi un Axolotl noir et un Axolotl blanc ne donnent pas forcément naissance à des petits Axolotls gris. On apprend aussi pourquoi la consanguinité peut être utile, ou au contraire mauvaise.
1. Vocabulaire
- ADN : programme de fabrication et de fonctionnement d'un organisme.
- chromosome : élément constitué d'ADN.
- allèle : une des deux versions d'un même gène, chacun se trouvant sur l'un des deux chromosomes de la paire.
- génotype : code génétique que porte l'Axolotl.
- phénotype : caractère visible d'un Axolotl (ex. : couleur).
- homozygote : qui possède deux allèles identiques sur un gène (l'un reçu du père, l’autre de la mère) ; par exemple, pour l’albinisme, on écrit a/a.
- hétérozygote : qui possède deux allèles différents sur un gène (l'un reçu du père, l’autre de la mère) ; par exemple, pour l’albinisme, on écrit A/a.
- gène dominant : qui s'exprime qu'il soit homozygote ou hétérozygote ; c'est le cas de la couleur Sauvage.
- gène récessif : qui ne s'exprime que s'il est homozygote ; c'est le cas des couleurs autres que Sauvage.
- gène porté : gène non exprimé ; par exemple, un Axolotl Albinos peut être porteur du gène Leucistique.
- gène exprimé : gène qui s'est exprimé ; par exemple, a/a (homozygote pour l'albinisme, l'Axolotl est bel et bien Albinos).
2. La théorie de base de la génétique
Tout ce qui compose un être physiologique est codé dans l'ADN. L'Axolotl a 28 chromosomes (soit 14 paires), tandis que l'Homme en a 46 (soit 23 paires). Chaque gène utilise une place particulière sur un chromosome.
Tout caractère héréditaire — par exemple, la couleur — d'un Axolotl est codé par un gène. La couleur de l'Axolotl dépend de cellules pigmentaires appelées « chromatophores » et des gènes qui les laissent ou non s'exprimer.
Chaque gène est composé de deux parties. Ces parties sont appelées « allèles », l'une étant donnée par la mère et l'autre par le père. Certaines ont besoin d'être à deux semblables pour s'exprimer : par exemple, un Axolotl a besoin de deux allèles (a) albinos, pour être albinos.
Lorsque le spermatophore du mâle — contenant 14 chromosomes — féconde l'œuf de la femelle — contenant également 14 chromosomes —, une fusion se produit pour former une cellule complète de 28 chromosomes, appelée « zygote ». Cette cellule est la première qui se multipliera pour devenir un Axolotl.
La progéniture se compose donc des gènes transmis par les deux parents, mais la variation génétique est telle que chaque nouvelle larve est génétiquement distincte de ses frères et sœurs et de ses parents.
Une toute dernière chose : certains gènes — en particulier pour ce qui concerne l'expression des couleurs — ont besoin d'être identiques pour s'exprimer : dans ce cas, on parle de gène « homozygote ». Par exemple, un Axolotl a besoin de deux gènes (a) albinos pour être Albinos.
Une fois que vous avez compris ces principes fondamentaux, vous êtes déjà bien armés, ![]() !
!
3. Les chromatophores
La variation de couleurs chez l'Axolotl est due aux « chromatophores », c'est-à-dire à des cellules de couleurs présentes dans la peau de l'Axolotl. Il en existe 3 sortes :
- les « mélanophores », composés d'eumélanine pigments bruns et noirs),
- les « xanthophores », composés de caroténoïdes et ptéridines (pigments jaunes et rouges),
- les « iridophores », composés de purines cristalisées, qui donnent une irisation brillante.
4. Les principaux gènes de couleur
Chaque Axolotl possède des gènes qui vont agir sur les chromatophores, en les laissant ou en les empêchant de s'exprimer, par exemple.
C'est comme dans le domaine de la photo ou de la peinture, lorsque l'on mélange différentes couleurs ensemble pour en obtenir de nouvelles : il faut du bleu et du jaune pour obtenir du vert mais, si l'on prend du vert et que l'on supprime ou filtre le bleu qu'il contient, il ne reste plus que du jaune. Eh bien, avec les chromatophores, c'est exactement la même chose, ![]() ! C'est grâce aux chromatophores et à la façon dont ils s'expriment (ou non) que l'on obtient les différentes couleurs des Axolotls.
! C'est grâce aux chromatophores et à la façon dont ils s'expriment (ou non) que l'on obtient les différentes couleurs des Axolotls.
Voici donc les différents gènes influençant les chromatophores :
| Nom | Gène a |
Gène ax |
Gène d |
Gène m |
Gène co |
Génotype complet |
| Sauvage | A- | AX- | D- | M- | CO-? | |
| Mélanique | A- | AX- | D- | mm | CO-? | m/m |
| Leucistique | A- | AX- | dd | M- | CO-? | d/d |
| Leucistique mélanique | A- | AX- | dd | mm | CO-? | d/d m/m |
| Albinos doré | aa | AX- | D- | M- | CO-? | a/a |
| Albinos axanthique | aa | axax | D- | M- | CO-? | a/a ax/ax |
| Albinos leucistique | aa | AX- | dd | M- | CO-? | a/a d/d |
| Albinos mélanique | aa | AX- | D- | mm | CO-? | a/a m/m |
| Hypomélanique?cuivre | A- | AX- | D- | M- | coco ? | co/co ? |
Albinisme (a) : incapacité de synthétiser la mélanine, agit donc sur les mélanophores mais les autres pigments ne sont pas affectés. L'axolotl n'aura pas de pigments foncés et il semblera avoir beaucoup de xanthophores (d'ou la couleur jaune !). Ce gène est récessif.
A/A est sans effet, individu non porteur
A/a est sans effet, individu porteur
a/a = Individu albinos
Axanthisme (ax) : absence d'iridophores et de xanthophores. Les mélanophores ne sont pas affectés donc l'animal sera gris foncé. Ce gène est récessif. Pas répandu.
AX/AX est sans effet, individu non porteur
AX/ax est sans effet, individu porteur
ax/ax = Individu axanthique
Leucistisme (d) : affecte la distribution des cellules pigmentaires qui ne peuvent pas migrer depuis la crête neurale sur le corps lors du développement de l'embryon. Bien que quelques tâches de mélanine puissent quand même apparaître immédiatement ou avec le temps, l'animal est blanc aux yeux noirs (les mélanophores qui pigmentent la rétine proviennent eux d'un diverticule du tube neural, non stoppés par le leucistisme). Ce gène est récessif.
D/D est sans effet, individu non porteur
D/d est sans effet, individu porteur
d/d = Individu leucistique
Mélanisme (m) : absence d'iridophores. L'axolotl aura plus de mélanoides et de xanthophores. Il n'aura aucun relfet irisé ni sur le corps ni dans les yeux. Ce gène est récessif.
M/M est sans effet, individu non porteur
M/m est sans effet, individu porteur
m/m = Individu mélanique
? Hypomélanisme (h) : réduction des mélanophores. L'axolotl aura une diminution de la pigmentation noire sur la peau, mais gardera un ton brunâtre et les yeux foncés. Les tons jaunes seront favorisés. Ce gène est récessif. Nous supposons que ce gène est à l'origine de la couleur biscuit ou cuivre chez l'axolotl, en comparant avec les connaissances d'autres espèces et les observations des éleveurs.
? H/H est sans effet, individu non porteur
? H/h est sans effet, individu porteur
? h/h = Individu hypomélanique
Il y a des gènes encore mal connus.
Ces gènes peuvent être actifs en même temps et promettre de belles couleurs.
Il est possible que cette liste s'étoffe avec le temps.
Les phénotypes, ou couleurs exprimées
Sauvage : Cette couleur est composée de nuances de brun avec taches noires, jaunes et brillantes. L'axolotl est sombre. Il n'est pas mélanique, ni albinos, ni axanthique en même temps. C'est la couleur de base de l'axolotl. Un axolotl sauvage peut être hétérozygote pour d'autres couleurs (porter d'autres gènes sans les exprimer).
Mélanique : Cette couleur est caractérisée par une absence de pigments brillants. L'aspect jaune est réduit, il y a plus de mélanophores et l'animal semble noir. Plus l'axolotl possède de mélanophores, et plus il sera sombre.
Un mélanique est homozygote pour m (m/m) mais peut être hétérozygote pour d'autres couleurs (porter d'autres gènes).
Leucistique : Cette couleur est caractérisée par un corps blanc ou rosé, tout en gardant les yeux noirs avec un cercle doré. La présence de quelques melanophores est possible sur la tête. En réalité d/d empêche les cellules pigmentaires de migrer sur le corps de l'animal.
Un leucistique est homozygote pour d (d/d) mais peut être hétérozygote pour d'autres couleurs (porter d'autres gènes).
Leucistique mélanique : Cette couleur est blanche comme un leucistique normal, cependant les yeux n'ont pas de cercle doré. L'animal n'a pas de pigments brillants, mais les autres cellules ne peuvent pas migrer sur le corps, d'ou l'apparence blanche. Un leucistique mélanique est homozygote pour d et pour M (d/d m/m).
Arlequin : Cette couleur est le corps de base d'un leucistique mais avec une forte présence de mélanophores. L'axolotl présente des tâches noires importantes ou larges et diffuses en particulier sur la tête et le long du voile dorsal. En réalité l'axolotl est leucistique, et le gène d/d qui empêche les cellules pigmentaires de migrer sur le corps de l'animal, en laisse quand même passer un certain nombre. Un arlequin sera génétiquement leucistique, donc homozygote pour d (d/d) et peut être hétérozygote pour d'autres couleurs (porter d'autres gènes).
Albinos : Cette couleur ne laisse aucun mélanophore se montrer, donc l'axolotl n'a visuellement aucun pigment noir. Il existe plusieurs types d'albinos.Un albinos est toujours homozygote pour a (a/a) mais peut aussi être hétérozygote ou homozygote pour d'autres couleurs.
Nous les détaillons ici :
Albinos doré : Cette couleur est jaune et dorée. Il s'agit d'un simple manque de mélanophores. Un gold est homozygote pour a (a/a)
Albinos axantique : Cette coulleur est blanche, mais l'animal peut se teinter en jaune avec l'âge en accumulant des riboflavines issu de l'alimentation. Il s'agit d'un manque de mélanophores de xantophores et d'iridophores. Un albinos axantique est homozygote pour a et ax (a/a ax/ax).
Albinos leucistique : Cette couleur est blanche. L'animal possède des pigments jaunes, mais le leucistisme les empêche de migrer sur le corps. Un albinos leucistique est homozygote pour d et a (d/d et a/a)
Albinos melanique : Cette couleur est blanc mat. C'est une combinaison entre mélanisme et albinisme qui supprimera tous les pigments ou les empêchera de se montrer, sauf un soupçon infime de xanthopores jaunes sur la tête et le dos. Un albinos melanique est homozygote pour m et a (m/m et a/a).
Cuivre : Cette couleur est composée de nuances de brun clair avec des taches brun foncées, jaunes cuivrées et brillantes mais peu ou pas de tâches très noires. L'axolotl n'est pas très sombre. Il n'est pas mélanique, ni albinos, ni axanthique en même temps. Un hypothèse serait qu'il s'agissent d'hypomélanisme (h) et donc l'individu hypomélanique est homozygote pour h (h/h). Dans tous les cas l'expérience des éleveurs montrent que le cuivre est une couleur récessive. Nous l'appèleront ici co (co/co).
Chimère : Ce n'est pas une couleur mais un concept d'individu fusionné, ne possédant pas le même code génétique partout dans son corps. Visuellement la chimère peut donc montrer deux couleurs différentes, l'une à droite et l'autre à gauche, et ce n'est qu'ainsi qu'on peut la reconnaitre. Pour qu'on la remarque eIle est par exemple moitié Albinos doré / moitié sauvage, ou moitié leucistique / moitié sauvage. Une chimère est un rare accident de la nature.
Cause et enchaînement détaillé des hasards provoquant une chimère naturelle in vivo :
1. Juste après la fécondation, des zygotes jumeaux se forment dans le même oeuf (on peut estimer 1% de chances dans une ponte)
2. Il faut que l'un des zygotes jumeaux migre contre l'autre dans un stade très primaire de développement (on peut estimer 1% de chance dans une ponte)
3. Il faut que les zygotes fusionnent pour n'en former qu'un seul composé des gènes de chacun (à ce stade là, l'embryon est juste un hamas de cellules !)
4. Il faut que cette "greffe" soit acceptée, avec l'axolotl on peut estimer que cela réussisse dans 100% des cas.
4. Il faut que le développement de l'embryon continue normalement, sans s'arrêter. (estimation 90 %)
5. Il faut que l'axolotl "moitié-moitié" naisse et grandisse (pourcentage variable entre selon la ponte et les conditions d'élevage, incalculable)
Les chances d'avoir une chimère seraient minimum de l'ordre de 1 pour cent mille (1/ 100'000), si l'on fait un grossier calcul.
Les capacités formidables de régénération de l'axolotl, et sa très bonne acceptation des greffes sont sans doute pour quelquechose dans le fait qu'on trouve un peu plus de chimères chez l'axolotl qu'ailleurs dans le règne animal.
Calculer les probabilités de couleurs dans les pontes
Pour appliquer la théorie de base, nous passons directement aux calculs. En effet, il est possible pour chaque axolotl et chaque ponte de faire des calculs statistiques, pour autant qu'on ait un maximum d'indications sur la lignée. Nous allons vous montrer comment.
Lors de la reproduction nous avons vu que chaque parent va donner l'un de ses deux allèles à ses petits. Dans ce cas chaque petit reçoit un gène de chaque parent. Pour facilité le calcul, nous avons établi un tableau comme il en est coutume. Nous voyons ainsi les gênes des parents et comment ils sont ditribués aux petits.
Chaque carré de petit (ceux qui sont moitié/moitié), équivaut à un quart de la ponte, soit 25%. En effet, en additionnant ces 4 carrés de 25% chacun, nous obtenons 100% c'est à dire la totalité de la ponte.
Rappelons que A veut dire gène non présent, et a gène présent en ce qui concerne l'albinisme.
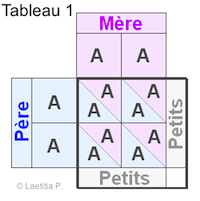 |
Exemple 1 En faisant le tableau, nous voyons que chacun des 4 carrés est semblable. |
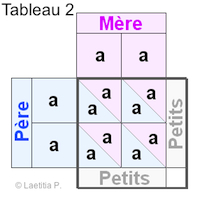 |
Exemple 2 En faisant le tableau, nous voyons que chacun des 4 carrés est pareil : |
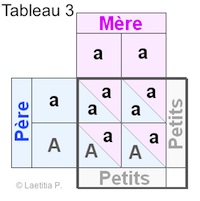 |
Exemple 3. Reproduction d'un couple d'axolotl, dont l'un est albinos et l'autre n'est que porteur. En faisant le tableau de croisements, nous voyons qu'il y a 2 sortes de carrés : |
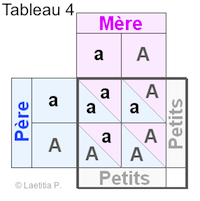 |
Exemple 4 En faisant le tableau de croisements, nous voyons qu'il y a 3 sortes de carrés : |
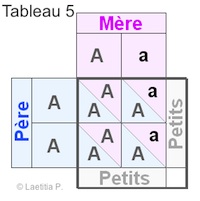 |
Exemple 5 50 % des petits sont A/a, donc non albinos mais porteurs. |
Bien sûr, les calculs se compliquent lorsque nous prenons en compte les gènes pouvant se combiner, mais pour l'exercice nous avons choisi de faire simple pour que vous compreniez le principe : tout peut se calculer lorsqu'on connait l'arbre généalogique ! Mais il reste des inconnues car on ne connait jamais tout d'un individu.
N'oublions pas mère nature !
Mère nature doit bien rire devant nos calculs savant pour essayer de deviner ce qu'elle fera naître ! En effet, malgré tous nos calculs, la nature ne respecte pas toujours les statistiques. Il y a des inconnues, des approximations et des surprises !
Un petit exemple pour l'homme : Selon la génétique, on a 50% de chances d'avoir un garçon, et 50% de chances d'avoir une fille. Pourtant des familles ont 3 gaçons et aucune fille !
Les axolotls des différents éleveurs produisent parfois une descendance dont le phénotype défie ce que l'éleveur sait de son génotype, parcequ'on ne sait jamais tout de la lignée d'un axolotl.
Lignée
Une lignée c'est les axolotls issus en ligne d'un ancêtre commun, considéré comme fondateur. (exemple de lignée : petits enfants - enfants - pères - grand père). Sachez qu'aucun gène ne se perd ! Ce qui veut dire que parmis les gênes de tous les petits enfants réunis, on retrouve tous les gènes du grand-père ! Et que si vous avez des mélaniques dans une ponte, c'est que quelque part dans les ancêtres du père et dans ceux de la mère, il y a au moins un individu mélanique. Etc pour chaque couleur et mutation, hormis pour les chimères (qui sont plutôt des accidents de la nature). Mais on ne connait jamais toute la lignée d'un axolotl, et la reproduction peut nous révéler de nombreuses surprises sur les gènes portés par les parents !
On ne sait jamais tout ce que porte un axolotl, car ses gènes peuvent venir de très loin en arrière.
Il est interressant de conserver trace des pontes, des ancêtres, etc. En effet, que ce soit une couleur ou une tarre génétique, tout peut se transmettre par le biais de la reproduction ! Mieux vaut donc pouvoir choisir quels individus vous allez éloigner de la reproduction, et pour quels individus vous souhaitez la favoriser.
Afin de mieux contrôler les lignées, certains éleveurs aiment isoler le couple qu'il souhaite se voir reproduire. C'est là que commence le travail de lignée.
Des gènes bons, et des gènes moins bons...
Il existe également des gènes et mutations moins bons, dangereux voir mortels. Ils se transmettent également lors de la reproduction. Tous les gènes abordés ci-dessous proviennet de l'étude de l'Ambystoma Genetic Stock Center et doivent être homozygotes pour s'exprimer.
- Anémie (an) : Les individus expriment une anémie temporaire à l'âge de 3-4 mois.
- Insuffisance cardiaque (c) : Le coeur se développe mais ne peut pas se contracter, la larve ne mange pas et meurt peut après l'éclosion.
- Absence d'oeil (e) : Les animaux n'ont pas deux (ou en partie), ils seraient stériles.
- Rétention d'eau (f) : L'axolotl fait une rétention d'eau (lymphe) déjà dans l'oeuf, tous ne sont pas viables.
- Branchies anormales (g) : développement anormal des branchies, suivit de la mort dans la semaine après éclosion. Les individus ont l'air plus foncés et plus jaunes que les autres.
- Mortelle après construction des mains (H) : Les larves se développement nomalement jusqu'à ce que les mains avant ayant 3-4 doigts. Certains axolotls montreront une courbure latérale du corps. Elles meurent rapidement.
- London Lethal (L) : Disproportion, tête très petite, mauvais développement des branchies. les larves grandissent moins vite et se nourrissent très peu. Ils meurent quelques semaines après éclosion.
- Microphtalmie (mi) : Leurs yeux sont petits, les pigments débordent sur la cornée, ils peuvent être asymétriques. Les larves mangent très peut et meurent rapidemment.
- Tête de broche (pi) : Grand développement de la tête. Les larves sont incapables de manger et meurent peu après l'éclosion. Les plus légèrement touchés peuvent survivre plus longtemps.
- Léthal (r) : Les larves meurent après quelques semaines.
- Orteils courts (s) : mutation qui implique les reins et les membres qui sont de taille réduite. Ils meurent d'une inssuffisance rénale aux environs d'un an.
- Spastique (sp) : N'arrivent pas à développer les mouvements de nage normale et droite.
- Congestion (st) : Couleur très rouge. La circulation du sang cesse partiellement ou totalement. Les larves meurent dans la semaine après éclosion, les plus téméraires survivront jusqu'à 4 mois.
- Twisted Gills (t) : les larves présentent une détérioration des branchies, leur développement rest retardé. Ils meurent deux semaines après éclosion environs car ils ne parviennent pas à se nourrir.
- Vasodilatation (v) La plupart des larves meurent au moment de l'éclosion. Les survivants seront plus petits.
- Létal (x) : Les branchies parraissent plus fragiles et se courbent vers l'avant aux extrémités. Les larves commencent à se nourrir mais meurent peu après.
10. La consanguinité
Bien qu'ils ne soient pas particulièrement sensibles à la consanguinité, le fait que des Axolotls d'une même lignée se reproduisent entre eux induit une fixation de certains gènes :
- certains pouvant servir d'outils dans l'expression de certains caractères visibles recherchés, comme la couleur, la longueur des branchies, etc. ;
- d'autres devant être écartés (comme ceux provoquant le caractère « boule », ou ceux conduisant à la létalité).
a. Quelques lumières de la part d'Emmanuel
Un grand merci pour ce mail explicatif,  .
.
« En tant que biologiste moléculaire de formation, la notion de consanguinité est un sujet qui m'intéresse même si, comme on le sait, la biologie n'est pas une science exacte et que personne ne peut affirmer de connaître la vérité sur le sujet. Il y a cependant pas mal de pistes qui vont dans le même sens pour affirmer que ce problème ne touche pas réellement les urodèles et, de ce fait, les Axolotls.
Il faut d'abord définir ce que l'on entend par « consanguinité ». Il existe pas mal de définitions, mais la notion de consanguinité est définie comme le fruit d’une reproduction entre deux individus apparentés.
Chez les organismes diploïdes (ayant des chromosomes groupés en paires), le coefficient de consanguinité est la probabilité pour que deux allèles (différentes versions d'un même gène) que possède un individu en un locus (emplacement sur un même chromosome) quelconque soient identiques par descendance. La consanguinité a globalement pour effet de réduire la valeur sélective (fitness) des individus/populations concerné(e)s, mais ceci de manière très variable. Elle augmente le risque de certaines maladies génétiques et d'occurrences de certains syndromes. Ces effets vont fortement dépendre des espèces ou populations considérées, et de la manière dont leur génotype (information portée par le génome d’un organisme) interagissent avec l’environnement.
Chez l'Homme, il est incontestable que la procréation consanguine concentre certaines caractéristiques génétiques défavorables. Pas étonnant, donc, que nous soyons amenés à faire le lien avec l’Animal : c’est typique de notre réaction anthropomorphique, selon laquelle on attribue des caractéristiques comportementales ou morphologiques humaines aux animaux.
Néanmoins, ça ne marche pas ainsi.
Il faut savoir que le fitness génétique qui est, par exemple, le nombre de descendants viables et fertiles que produit en moyenne chaque individu de ce génotype à la génération suivante, aussi connu sous le nom de valeur sélective qui est une mesure de la sélection naturelle, a tendance à décroître avec l’évolution. Je ne parle pas de l’évolution de l’Homme, mais du protozoaire par exemple vers les mammifères et en particulier les animaux dits supérieurs comme les primates (Homme inclus, bien entendu).
Aussi le pool génétique et les capacités adaptives ont-ils tendance à diminuer avec l’évolution, et les problèmes liés à la consanguinité seront-ils d’autant plus importants au sommet de l’évolution qu'à sa base.
Même parmi les mammifères, il y a de grandes différences : certains animaux ne semblent pas y être sensibles, comme par exemple les chèvres qui ont pu coloniser certaines îles à partir d’un très petit nombre d’individus (libérés par les marins, à l’époque), et pourtant leur population est saine. Il en va de même pour le rat, le chat ou le lapin (en Australie), le cheval et pas mal d’autres espèces. Les cas sont nombreux. Par contre, certaines espèces sont très affectées et sont en bout de course, comme le guépard, le bison d’Europe, l’ours blanc, qui n’ont plus la capacité de renforcer leur population . Ils vont inéluctablement vers l’extinction, car ils sont en bout de course génétique.
Les urodèles, et donc l'Axolotl, sont dans la base de l'évolution, donc à un stade primitif. Leur fitness génétique n’est pas menacé, mais beaucoup d’autres menaces planent malheureusement sur eux, mais ceci n’est pas l’objet de la question. Pour la petite histoire, la quasi-totalité des Axolotls détenus en Europe par les privés ou les laboratoires provient de 6 animaux capturés au début du siècle dernier et placés au Jardin des Plantes à Paris. À partir de ce peu d’animaux, une énorme population s’est constituée, il y a eu des centaines de générations, des milliers de croisements, nous avons réalisé une multitude de variations de couleurs et de formes et des sélections successives.
Si l'Axolotl était touché par la consanguinité, cela fait bien longtemps qu’il aurait disparu d’Europe car, à ma connaissance, une seule autre importation, très probablement illicite, aurait été faite par des Allemands il y a une dizaine d’années.
Par ailleurs, les urodèles, et bien entendu l'Axolotl, ont la capacité de régénérer leurs membres et organes non vitaux. Ceci est fortement étudié, comme par exemple à l’Université de Genève pour une application à l’Homme. Il y a cependant encore beaucoup à apprendre. Cette capacité a elle aussi été perdue dans l'évolution et s'arrête aux urodèles (en dehors de la queue du lézard).
Ce phénomène est complexe, mais trouve une explication au niveau moléculaire : un mécanisme de réparation hyper performant qui répare également les cellules et même le noyau. Lors de la méiose, il va y avoir division de la cellule aboutissant aux cellules sexuelles (ovules et spermatozoïdes), dans laquelle les paires de chromosomes du noyau se séparent pour former des gamètes, qui s'uniront avec un gamète de l'autre sexe pour former un œuf.
Cette opération est très complexe et peut former un grand nombre de mésappariements qui vont créer des erreurs dans le génotype du futur individu (tares, maladies, malformations etc.) qui est d’autant plus augmenté par la procréation consanguine. Les urodèles sont privilégiés du fait de leur capacité intrinsèque à réparer ces erreurs, nous n’avons pas cette faculté et cette chance.
On peut d'ailleurs remarquer le peu de larves mal formées sur la quantité d’œufs pondus.
Par ailleurs, de vieux individus sont toujours capables de se reproduire et d’avoir une descendance viable, ce qui n’est pas le cas de l’espèce humaine.
Tout ceci pour dire que la consanguinité, à ma connaissance, n’a jamais été observée chez l'Axolotl et qu’elle fait plus partie d’un mythe que d'une réalité scientifique. »
b. Article scientifique
Pour celles et ceux qui lisent l'anglais, voici un article scientifique démontrant que, chez les Urodèles, les malformations sont liées à des facteurs extérieurs (comme la pollution), et non pas à la consanguinité : Amphibian malformations and inbreeding.
Date de dernière mise à jour : mardi 02 mars 2021